LE CRÉPUSCULE DES DINOSAURES
L’extinction de masse qui marqua la fin du Crétacé semble stimuler l’imagination des scientifiques. Des savants parfaitement respectables qui s’enorgueillissent de leur prudence quand ils travaillent dans leur propre spécialité, se permettent les plus fantastiques envols de leur imagination quand ils en viennent à se pencher sur le mystère de l’assassin de la fin du Crétacé. J’ai tout un dossier sur les « solutions » publiées. Il contient un certain nombre de suggestions : "les dinosaures ont disparu parce que le temps devint trop chaud" ; "parce que le temps devint trop froid“ ; "parce que le temps devint trop sec" ; "parce que le temps devint trop humide" ; "parce que le temps devint trop chaud en été et trop froid en hiver" ; "parce que le terrain devint trop vallonné" ; "parce qu’on vit apparaître de nouvelles plantes qui empoisonnèrent tous les dinosaures " ; "parce qu’on vit apparaître de nouveaux insectes qui répandirent des maladies mortelles" ; "parce que de nouvelles espèces de mammifères mangèrent les œufs de dinosaures" ; "parce qu’une météorite géante s’écrasa sur la Terre" ; "parce qu’une supernova explosa près de la Terre" ; "parce que des rayons cosmiques bombardèrent la Terre" ; ou “parce que des volcans géants entrèrent en éruption tout autour de la Terre“.
Il m’a toujours semblé curieux que des savants, par ailleurs assez austères, puissent sauter sur des conclusions étonnantes quant à la disparition des dinosaures. Peut-être, comme Zorba le Grec nous l’a enseigné, savants et non-savants sont-ils enclins à croire à des solutions tirées par les cheveux parce que nous avons tous besoin d’un peu de folie. Les événements dont il s’agit n’ont pas eu de témoins directs et furent d’une dimension exceptionnelle, supérieure même à la vie, différents de tout ce que nous pouvons voir dans notre monde moderne. Par voie de conséquence, nous sommes fascinés, attirés par l’idée que les mystères à très grande échelle réclament des solutions tout aussi fantastiques, des solutions totalement différentes des événements quotidiens que nous vivons tous les jours.
Je suis tout à fait convaincu que tous les grands mystères - les pyramides mayas, Stonehenge, les têtes de l’Île de Pâques et l’extinction des dinosaures peuvent être résolus. En fait, je crois que la plupart d’entre eux ont été résolus depuis longtemps. Mais la solution est en général si évidente, si peu fantastique, que sa banalité nous secoue. Quelle peut être la solution banale et évidente de la grande extinction du Crétacé ? Construisons la preuve morceau par morceau, avant d’annoncer le nom du criminel.

L’analyse des indices
Tenter de découvrir le coupable de l’extinction de masse commence par la solution du problème de base de la criminologie : la fiabilité des indices. Les savants ont parfois espéré que les carcasses de dinosaures les conduiraient à découvrir l’agent létal. Dans les années 1920, un paléontologue conclut que les dinosaures à bec-de-canard étaient morts empoisonnés, car leurs squelettes étaient tordus dans ce qui semblaient être des postures d’agonie. Mais il s’avéra que ces squelettes étaient tordus - le cou vrillé vers le haut et l’arrière - parce que c’est la position normale d’un cadavre, les muscles et tendons du cou se contractant après la mort. En réalité, les squelettes fossiles livrent rarement, sinon jamais, des indices sur la cause de la mort que ce soit à la fin du Crétacé ou à toute autre époque. Des coupes dans les os de dinosaures ne montrent habituellement aucun signe pathologique. Mais même cela ne permet pas de conclure parce qu’il est rare que les maladies mortelles laissent des marques dans les os.
Les seuls indices pour découvrir le coupable sont donc ceux des circonstances, les paramètres concernant le lieu du crime. Le problème avec de telles preuves est de séparer les faits probants de ceux qui ne prouvent rien. Un procédé assez commun, adopté par tous ceux qui cherchent à résoudre le mystère, consiste à comparer tous les détails des circonstances de la mort des dinosaures en Amérique du Nord que nous connaissons avec tous les morceaux d’informations en provenance des autres endroits du monde. Dans ce cas, l’hypothèse suppose qu’il y a eu quelque rupture dans l’habitat qui a provoqué la disparition des dinosaures. De ce fait, il faut se concentrer sur ce qui a changé à la fin du Crétacé. Le défaut majeur de cette approche est que, sur terre, les divers habitats ne sont jamais stables ; ils changent en permanence. Si nous cherchons la preuve d’un changement dans l’environnement, n’importe où dans l’histoire géologique, nous sommes certains de le trouver.
Pour prendre un exemple des problèmes inhérents à cette méthode de recherche des circonstances, les paléontologues qui s’intéressent aux plantes ont découvert que les hivers étaient devenus plus froids a la fin du Crétacé. De ce fait, de nombreux géologues ont conclu que les dinosaures étaient morts de froid. Mais cette conclusion est prématurée. Il est possible que ce changement climatique soit innocent, un événement sans importance contemporain du véritable agent de la disparition des dinosaures. On ne peut le désigner comme coupable seulement parce qu’il était présent sur les lieux du crime.
La première étape pour évaluer l’importance d’une preuve dans ce problème de l’extinction de masse est malheureusement celle qu’on ignore le plus souvent : la recherche d’un modèle qui se répète au long du temps. Bien trop souvent, l’extinction des dinosaures est perçue comme une brutalité écologique isolée, une Saint-Barthélemy évolutive infligée aux habitants des plaines et forêts du Crétacé. Si l’extinction avait été un événement unique et jamais répété, alors il serait presque impossible de trier les coïncidences sans intérêt des preuves désignant le coupable. Mais si ce coupable est un assassin récidiviste qui a frappé l’écosystème à plusieurs reprises dans les temps géologiques, nous aurions une chance supplémentaire de pouvoir éliminer les preuves qui n’en sont pas. Des attaques répétées par un même agent, dans des circonstances variées, doivent révéler un modus operandi qui serait la marque caractéristique du criminel.
La première étape pour résoudre le mystère de l’extinction de masse de la fin du Crétacé consiste donc à se poser des questions. Y a-t-il eu d’autres extinctions de masse ? La réponse est "Oui". Les extinctions de masse ont frappé à la fin du Permien, quand les gorgonopsiens des familles tatariennes disparurent en même temps que leurs proies, les dicynodontes ; à la fin du Trias quand les gros porteurs de défenses sur terre et les lézards marins dans les océans furent balayés ; à la fin du Jurassique quand de nombreuses (mais pas, toutes) lignées dinosauriennes disparurent. En tout, les documents fossiles indiquent huit extinctions de masse brutales qui frappèrent les gros vertébrés terrestres. La plus récente est arrivée voici dix mille ans quand la plupart des espèces géantes de mammifères – les mammouths, les mastodontes, les chameaux géants, les tigres à dents de sabre et quelques autres - périrent.

L’étape suivante consiste à se demander si ces extinctions de masse suivent un modèle cohérent. La réponse est encore affirmative ; la loi de Cuvier sur la simultanéité sur terre et en mer s’applique. À chaque fois que l’écosystème terrestre a subi une extinction de masse, l’écosystème marin a également souffert. Quand les dinosaures furent balayés a la fin du Crétacé, les grands lézards marins et les plésiosaures à cou serpentiforme disparurent aussi, en même temps qu’une flopée d’invertébrés, grands et petits, des huîtres coralliennes aux Ammonites et au plancton microscopique. La même simultanéité terre-mer a marqué le désastre du Tatarien, les extinctions du Jurassique et toutes les autres Grandes Extinctions, y compris celle qui frappa la classe des mammifères à la fin de l’Éocène. La nature double, terrestre et marine, de ces extinctions élimine automatiquement de nombreux criminels possibles. La disparition du Crétacé par exemple ne peut s’expliquer par l’évolution de plantes vénéneuses car les animaux marins n’auraient pas été affectés par leur toxicité.

L’étape suivante fera remarquer que les extinctions ont frappé la terre et la mer au même moment mais de manière différente. Tout le système marin a souffert. L’extinction des animaux minuscules, planctoniques, a été aussi complète que celle des serpents marins géants. Les extinctions ont éliminé la plupart des grands animaux, mais les habitants des milieux dulçaquicoles n’ont pas été touchés. Crocodiles, alligators et tortues d’eau douce n’ont pratiquement pas change après le Crétacé. Une autre différence entre animaux terrestres et animaux marins est que les petits animaux terrestres n’ont pas souffert autant que les gros. Les grands dinosaures du Crétacé ont complètement disparu, mais la plupart des familles de lézards et de mammifères ont traversé le désastre sans perdre leur pouvoir évolutif. Finalement - et c’est un point largement ignoré par la plupart des scientifiques - les extinctions de masse sur terre ont frappé plus fort les groupes de grands animaux les plus dynamiques, ceux qui évoluaient le plus rapidement, les familles qui produisaient le plus d’espèces et celles qui montraient les capacités adaptatives les plus élevées. Ce sont les groupes même où nous possédons les preuves les plus fortes d’endothermie : les proto-mammifères, les crocodiles pourpres, les ptérodactyles, les dinosaures et les grands mammifères de l’Éocène.
Les extinctions terrestres possèdent une autre particularité. En moyenne, les plantes ne souffrent pas autant que les herbivores. Les plantes de la fin du Crétacé ont souffert de quelques disparitions, mais rien de comparable à la disparition totale des dinosaures herbivores. Cette relative immunité des plantes reste vraie pour les autres extinctions, précédentes et suivantes.
À ce point, le modus operandi de l’assassin lors des grandes extinctions commence à être connu. Le coupable : (1) frappe sur terre et en mer au même moment ; (2) frappe plus durement les familles terrestres de grands animaux évoluant vite ; (3) frappe moins fort les petits animaux terrestres ; (4) laisse intacts les grands animaux à sang froid ; (5) ne frappe pas les animaux dulçaquicoles - la plupart de ces animaux sont à sang froid et donc le critère (4) s’applique ; (6) frappe les herbivores plus sévèrement que les plantes.
Un agent connu peut-il opérer selon ce modèle ? Aujourd’hui, la plupart des scientifiques croient que oui. La presse scientifique est pleine d’hypothèses nouvelles donnant la solution au problème, dont l’une est complètement extraterrestre. L’Étoile de Mort au cycle de 26 millions d’années serait un corps céleste géant qui frappe l’écosystème dans son ensemble à chaque fois qu’elle passe près de la terre. De telles théories ne sont pas neuves.
La théorie d’Alvarez
Un impact de météorite a déjà été proposé comme cause d’extinction voici quelques années. On n’en avait aucune preuve jusqu’à ce que Walter Alvarez, un chimiste spécialisé dans la microanalyse d’éléments rares, découvre la désormais fameuse couche d’iridium. L’iridium est un "métal noble", semblable au platine, mais plus dense, et comme le platine et l’or il ne se lie pas facilement avec d’autres éléments. Selon la théorie astrochimique classique, l’iridium est extrêmement rare à la surface de la Terre, mais très abondant dans les corps célestes, tels que les météorites, les astéroïdes et les étoiles mortes.
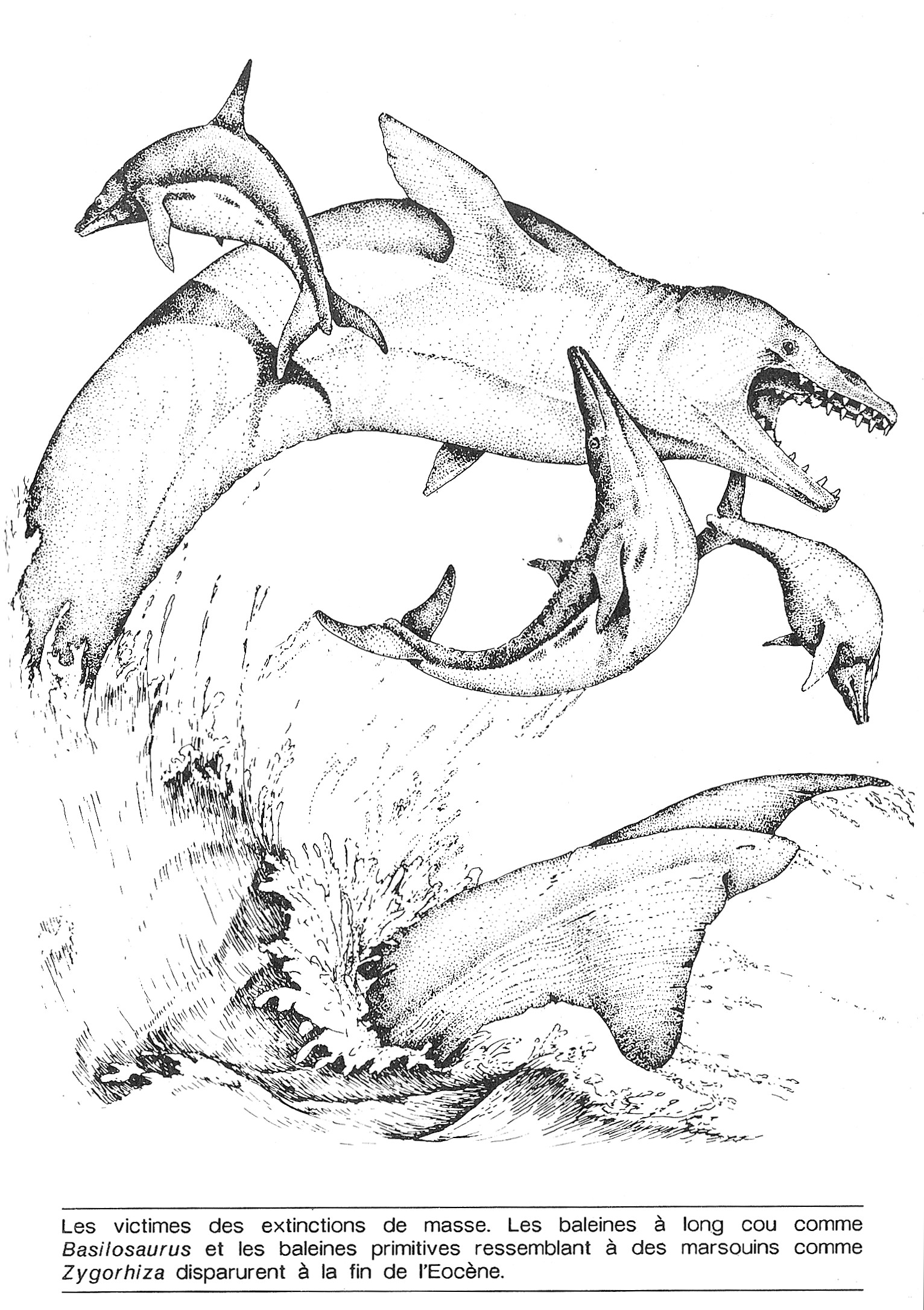
Alvarez n’avait pas commencé dès l’abord à chercher de l’iridium. Le but initial de sa recherche sur le Crétacé était routinier : il cherchait les moyens d’identifier la dernière couche sédimentaire déposée immédiatement après la fin du Crétacé. Si cette couche pouvait être identifiée, les géologues pourraient utiliser cet horizon comme modèle pour comparer la suite des événements géologiques tout autour de la planète. Ce type de marqueur a des applications intéressantes, car il est en général difficile de dater les couches en Europe par rapport aux couches américaines. Parfois, les couches de sédiments comportent des marques chimiques distinctives sur des millions de kilomètres carrés car l’activité volcanique peut étendre des nuages de fines particules de poussière au-dessus des océans du monde entier, qui déposent des concentrations uniques de minéraux dans la boue du fond des mers.
Alvarez utilisa des détecteurs ultra-sensibles sur les sédiments marins de Gubbio, en Italie, où une séquence exceptionnellement bonne de couches a été déposée à la fin du Crétacé. Un brutal changement des planctons fossiles marque la couche où les paléontologues placent la fin du Crétacé. À ce point exact, Alvarez tomba sur un marqueur géochimique frappant, avec des implications inattendues, une fine zone riche en iridium.
Alvarez et ses collègues annoncèrent leur découverte et étonnèrent la communauté paléontologique avec leur conclusion : une météorite géante avait frappé de plein fouet le monde des dinosaures. L’idée principale était qu’une météorite (ou un astéroïde) de grande taille, frappant la Terre à la fin du Crétacé, avait recouvert une immense surface de la planète d’une fine couche d’iridium parce que l’explosion de la masse céleste avait libéré d’énormes nuages chargés de poussière riche en iridium. Ces nuages de poussière se déposèrent ensuite sur la surface du globe et se mêlèrent au sol.
La théorie d’Alvarez en convertit beaucoup, essentiellement parce qu’une collision avec un corps céleste géant pourrait expliquer partiellement la sélectivité des grandes extinctions. Quand la masse de la météorite frappa la Terre, les nuages de poussière qui en résultèrent obscurcirent le ciel, cachant les rayons du soleil. Les plantes disparurent, puis les herbivores et finalement les carnivores lorsque la température chuta sous le parapluie noir des nuages. Les créatures à sang froid purent se cacher dans leurs terriers et attendre que la poussière se dépose, car leur métabolisme leur permettait de jeûner longtemps sans dommages. Ainsi, pouvait-on expliquer facilement que les tortues et les crocodiles n’aient pas été affectés. Les petits animaux pouvaient subir une telle extinction (quelques familles de mammifères disparurent), mais ils possèdent en général des terriers où se cacher et supporter les conséquences du froid. Et comme de nombreux petits animaux sont omnivores, ils auraient pu survivre en se nourrissant des dépouilles des dinosaures qui avaient succombé. De nombreuses espèces de plantes ont pu survivre dans un état de dormance – graines, spores, bulbes et tubercules - ce qui expliquerait le peu d’effet sur le monde végétal. Pour ceux d’entre nous qui sont convaincus que les dinosaures étaient endothermes, le grand nuage de poussière pourrait expliquer leur disparition. Leur métabolisme élevé et leur grande taille les rendaient plus vulnérables, car ils ne pouvaient échapper au désastre. Même la répétition des extinctions de masse pouvait être expliquée. Une comète pourrait suivre un cycle régulier de collisions avec la terre, une trajectoire qui permettrait aux rencontres de se renouveler chaque fois que les orbites de la Terre et de la comète se croiseraient. Une analyse mathématique publiée en 1983 affirmait que de telles extinctions frappaient régulièrement, tous les 26 millions d’années et donc la cause a même reçu un nom, l’Étoile de Mort au cycle de 26 millions d’années.
Je crois qu’il y a un cycle d’extinctions. Je ne crois pas à la théorie d’un choc venu du Cosmos. Un astronome de mes amis à Boulder m’a défié à ce propos. Je me fais l’avocat d’une large variété d’hérésies sur les dinosaures, aussi pourquoi ne puis-je accepter la théorie de leur extinction basée sur le choc d’une comète et fondée sur la couche d’iridium ? Ma défense est simple. Je défends les hérésies quand elles collent mieux aux faits que l’orthodoxie. La théorie de la météorite ne colle pas aux faits sur un point capital. Elle prétend que les extinctions étaient brutales, catastrophiques. Tous les dinosaures sont supposés avoir disparu en quelques dizaines d’années, ou approximativement. Mais depuis un certain temps, la paléobotanique orthodoxe a affirmé que les extinctions se sont étalées sur des dizaines de milliers d’années, ou plus encore. Et sans nul doute, l’orthodoxie a raison cette fois-ci. Les paléontologues travaillant au Montana affirment qu’ils observent une extinction graduelle des dinosaures et des mammifères du Crétacé et l’apparition graduelle de nouveaux groupes de mammifères qui vont dominer le monde à l’époque suivante. En fait, à de rares exceptions près (et Dale Russell du Musée National du Canada est la principale) les paléontologues sont tous d’accord. Les dernières extinctions n’étaient pas une boucherie qui a duré un petit week-end mais un long processus qui a demandé des milliers, voire même des millions d’années.
L’extinction graduelle
Ce dont nous avons besoin ici est une analyse soigneuse, couche par couche, des faunes fossiles. D’une telle analyse, nous pourrions établir un calendrier précis des extinctions. Si la dernière dynastie des dinosaures a commencé à décliner quelques millions d’années avant la couche d’iridium, la théorie de la collision cosmique perd toute validité. Peut-on établir un tel calendrier ? Malheureusement, la manière la plus simple et la plus populaire est aussi la plus dangereuse : il s’agit de compter les genres dans les échantillons de fossiles que nous avons. Il est impossible de connaître tous les genres de dinosaures qui vivaient à la même époque. Il est simplement, possible d’en identifier une fraction car beaucoup étaient si rares qu’ils ont eu peu de chances d’être seulement fossilisés. Plus nous découvrons de squelettes, plus nous pouvons trouver de genres. La seule façon correcte de comparer les quantités de genres est de juxtaposer les formations qui contiennent le même nombre de spécimens identifiables. La plus ancienne des trois formations de la fin du Crétacé en Alberta, celle de Judith River, a fourni plusieurs centaines de squelettes ; la suivante, celle de Scollard, n’en a fourni que quarante. De toute évidence, il y a une chute du nombre de genres entre les deux formations, mais cela peut être simplement dû à l’échantillonnage plus restreint de la Formation de Scollard.
Dale Russell a utilisé une procédure mathématique appelée "analyse de raréfaction" pour corriger ces chiffres. Cette analyse détermine combien de genres auraient été trouvés dans chaque formation si le nombre de squelettes avait été identique. Il en a conclu que le fossé entre Judith River et Scollard n’apparaîtrait pas si large si le même nombre de spécimens était disponible dans chaque formation. De cela, Russell a déduit qu’il n’y avait pas de crise chez les dinosaures à l’époque de Scollard. Il a insisté sur le fait qu’il n’y a aucune diminution significative du nombre de genres de dinosaures jusqu’à la fin de la dernière formation, celle d’Edmonton-Hell Creek où ils disparaissent tous en un instant.
Peut-on trouver ici une réponse correcte ? Les extinctions avaient-elles commencé bien avant que la couche d’iridium ait été déposée où arrivèrent-elles brutalement, au moment précis de la collision supposée ? Pour répondre à cette question, il est important de se rappeler que genres et espèces de dinosaures ont disparu tout au long du Crétacé, et même tout au long du Mésozoïque. Ce qui rend les extinctions de la fin du Crétacé si particulières est qu’aucune espèce n’apparaît pour remplacer celles qui disparaissent. D’une certaine manière, c’est le point capital des extinctions de masse : le taux des extinctions surpasse la production de nouvelles espèces, de telle sorte que des groupes entiers se trouvent entièrement vidés d’espèces. Mais Russell a raison en estimant qu’il devait y avoir bien plus de dinosaures que nous le savons à la fin de cette ère. De nouvelles espèces et de nouveaux genres ont sans nul doute continué à évoluer jusqu’à la fin de la période d’Edmonton-Hell Creek. Et, il a tort d’affirmer que le monde des dinosaures n’a subi aucun dommage jusqu’à ces tout derniers jours. Un très important composant de l’écosystème déclinait déjà dangereusement dès l’époque de Scollard.
Le paramètre qui déclinait dangereusement était l’égalité, que les écologistes appellent aussi l’équitabilité. Quand les écosystèmes sont bien portants et bien protégés de l’extinction, aucun genre ne domine. Il y a des genres presque aussi abondants dans chaque catégorie écologique : plusieurs herbivores, plusieurs carnivores, et ainsi de suite. L’époque de la Judith River était exactement comme cela. Aucun genre de dinosaure n’était vraiment dominant et les rôles principaux étaient répartis. Trois gros hadrosaures étaient communs : Corythosaurus, Lambeosaurus, Prosaurolophus et les dinosaures cornus étaient représentés par trois genres : Centrosaurus, Chasmosaurus et Styracosaurus. Une telle uniformité s’exprime en termes mathématiques par ce que l’on appelle l’Indice de Simpson. Sur l’échelle de Simpson, la faune de Judith River atteint 3,2.
L’Indice de Simpson a été élaboré pour répondre à une question implicite : quelle est la probabilité de trouver deux individus du même genre dans un échantillon si les animaux sont rencontrés au hasard ? Si la faune dans une aire donnée est assez uniforme, avec plusieurs genres assez communs, deux individus du même genre ne se rencontreront pas très souvent. De ce fait, une valeur élevée de l’indice de Simpson indique une faible probabilité de rencontrer deux individus du même genre dans l’échantillon et indique une grande uniformité. L’Indice de Simpson se calcule facilement : (1) prendre les espèces les plus communes : (2) prendre leur pourcentage dans la population totale, par exemple 1/3 du total ; (3) élever cette fraction au carré et le résultat est la probabilité de rencontrer cette espèce deux fois dans l’échantillonnage. Ainsi, si une espèce représente 1/3 de la faune totale, la probabilité de rencontrer deux animaux du même genre est (1/3)2, c’est-à-dire 1/9 ; (4) répéter l’opération pour toutes les espèces ; (5) ajouter toutes les probabilités et diviser un par le total pour avoir l’indice. Par exemple, supposons quatre genres de dinosaures qui représentent 1/3, 1/4, 1/4 et 1/6 du total. Leurs probabilités de rencontre seront de 1/9, 1/16, 1/16 et 1/36. En convertissant les fractions en nombres décimaux, ces probabilités seront de 0,11, 0,06, 0,06 et 0,03. Leur total vaut 0,26. Un divisé par 0,6 donne 3,8 et 3,8 est l’Indice de Simpson. À l’opposé, si une espèce représente 99.99 % de la faune, alors la probabilité sera de 0,999 et l’Indice de Simpson sera proche de 1.
Les dinosaures de la Formation de Judith River bénéficiaient d’un écosystème riche et uniforme, l’un des plus uniformes que l’on connaisse. Mais dans la couche suivante, celle de Scollard, cette uniformité disparaît. Un seul genre d’hadrosaure, Saurolophus, représente 75 % des gros spécimens et les autres sont assez rares. L’Indice de Simpson pour cette formation tombe à 1,4, ce qui est un score assez bas. Quelque chose est en train de se produire, les nouvelles espèces ne développent pas assez vite des adaptations leur permettant de prendre leur part de l’écosystème. Le déséquilibre se poursuit dans la formation suivante, celle d’Edmonton-Hell Creek, pour laquelle l’Indice de Simpson est de 1,3. Un seul genre, Triceratops, représente 70 a 80 % des découvertes de grands dinosaures. Cette situation inégalitaire dura deux millions d’années avant l’extinction finale.

Je devins sensible au concept d’uniformité dans l’évolution des dinosaures quand j’étais étudiant. Il était clair que, même si une collision avec une météorite avait détruit les derniers dinosaures, toutes les dynasties avaient été fortement affaiblies par d’autres causes. Je me mis à vérifier les autres extinctions de masse pour déterminer s’il y avait eu également une modification de l’uniformité avant la disparition finale. J’ai passé trois ans à accumuler des preuves dans les musées, en Afrique, en Europe et aux États-Unis. Après avoir compté et mesuré des milliers de crânes et de squelettes, la façon d’opérer du tueur vint en point de mire. Toutes les extinctions bien documentées suivaient le modèle de la fin du Crétacé. Bien avant l’extinction finale, l’uniformité se délitait. Les gorgonopsiens à dents de sabre de l’Époque Tatarienne en sont un parfait exemple. Leur écosystème montre les trois étapes typiques : un temps où la faune est riche ; une dégradation de l’uniformité s’ensuit ; et la chute, quand les gorgonopsiens disparaissent entièrement.
Le modèle historique que suivent les extinctions de masse ne corrobore absolument pas la théorie d’une destruction brutale des faunes par une Étoile de la Mort en quelques années. Mais alors quelle est la signification de la couche d’iridium ? Je n’ai aucune certitude. La sédimentation s’était beaucoup ralentie à la toute fin du Crétacé quand cette couche s’est déposée. Quelques géologues ont donc suggéré que les volcans avaient produit l’iridium qui s’était concentré parce qu’il était mélangé à des quantités inhabituellement faibles de sédiments. D’un autre côté, Erle Kauffman, un paléontologue de Boulder spécialisé dans le Crétacé, croit qu’un objet céleste a réellement frappé la Terre. Mais il pense que ce choc fut le coup de grâce porté à un écosystème mourant et souffrant de nombreux problèmes. Si j’en crois cette hypothèse, les dinosaures allaient diminuant bien avant la collision, mais le choc céleste mit un point final à un système moribond.
La solution ?
En toute hypothèse, l’histoire montre que les collisions célestes ne peuvent être les principaux coupables dans la disparition des écosystèmes. Tout au plus, ce sont des accessoires. Mais cela laisse la principale question sans réponse. Qu’est-ce qui détruit l’uniformité d’un écosystème ? Si je cherche une réponse, je suis étonné de voir le peu de cas que l’on fait de la vieille théorie des mers peu profondes. La meilleure réponse pour la disparition des animaux marins est que leurs terrains de chasse favoris disparurent quand les mers chaudes, peu profondes, se retirèrent des masses continentales. De même, la meilleure solution à la disparition des grands prédateurs marins est que la surface des océans devint plus froide et se mélangea mieux aux eaux profondes lorsque les mers se retirèrent. Ces changements dans le niveau des océans sont parfaitement documentés. Il n’est nul besoin d’une hypothèse extraterrestre pour cette extinction quand nous avons une excellente explication sur terre.
Cette théorie parfaitement établie du retrait des océans ne permet pas malgré tout d’établir comment ces modifications ont pu affecter des animaux terrestres de grande taille et actifs comme les gorgonopsiens du Tatarien ou les dinosaures du Crétacé. Les climats se refroidissent un peu sur terre quand les mers peu profondes se retirent parce que ces mers agissent comme des régulateurs thermiques. Mais les grands animaux très actifs sont en général plus résistants au froid que les plus petits ; pourtant, les extinctions de masse les frappent plus durement. Il doit y avoir autre chose qu’une raison climatique pour détruire ces géants vigoureux.

Le coupable le plus vraisemblable était une cause si banale et si terrestre qu’elle manque totalement de charme en face des collisions cosmiques. Et ce coupable a été clairement décrit dès 1925 par le grand paléontologue Henry Fairfield Osborn.
Observons la séquence historique qui se déroule sur terre lors d’une extinction de masse. Les mers peu profondes se retirent et les masses continentales qui se trouvaient sous l’eau s’exondent ; les régions qui se trouvaient séparées sont désormais reliées par des ponts continentaux ou des chapelets d’îles. Au même moment, la surrection des montagnes s’affaiblit et de moins en moins d’obstacles séparent les régions du monde. Bien entendu, de tels changements demandent des millions d’années. Mais le résultat le plus clair est un écosystème plus homogène où les espèces peuvent passer plus facilement d’un continent à l’autre et d’une extrémité de continent à une autre extrémité. Des échanges intercontinentaux ainsi facilités peuvent être observés précisément à la fin du Crétacé. Jusqu’à la fin du Crétacé, la Mongolie possédait une faune totalement différente de celle de l’Amérique du Nord. On rencontrait sur les hauts plateaux de l’Asie des mammifères très évolués et des dinosaures de la famille des protocératopsidés que l’on ne trouvait pas dans l’Alberta, le Montana et le Wyoming. Mais, tout à la fin du Crétacé, on voit apparaître les mammifères et les dinosaures asiatiques en Amérique du Nord. Ces émigrants n’ont pu passer que par le pont continental du Detroit de Bering, là où le nord-est de l’Asie touche l’Alaska.

Les autres époques de crises que l’on a pu bien étudier montrent le même symptôme, un accroissement des échanges d’espèces entre continents. Les grands mammifères terrestres furent sévèrement frappes par les extinctions au début de l’Éocène, époque où il y eut un échange extrêmement actif d’espèces entre l’Europe, l’Asie et l’Amérique du Nord. De tels échanges peuvent-ils conduire a l’extinction ?
La réponse est claire : c’est plus que possible. Un des dogmes inamovibles de la zoogéographie est que les pires conséquences sont possibles quand des espèces étrangères arrivent dans une région. Chaque espèce de reptile, d’oiseau ou de mammifère transporte avec elle son chargement de parasites et de maladies. La plupart de ces organismes étrangers ne trouveront pas d’ennemis sur place pour les contrôler et ils apportent la mort avec eux. Les pires épidémies qui ont balayé les hommes comme les animaux domestiques sont venues de l’introduction d’espèces étrangères (allogènes). La peste noire est venue de l’Asie avec les rats avant de balayer l’Europe. La peste bovine, maladie du bétail asiatique, débarqua en Afrique quand Lord Kitchener utilisa des bovins indiens pour tirer ses canons le long du Nil et déloger Marchand de Fachoda à la fin des années 1880. Lâchée au milieu des bovins africains, la peste bovine provoqua la mort des antilopes et massacra des millions de ruminants, du Soudan au Cap. Aujourd’hui encore, après un siècle de tentatives pour la contrôler, les gardes-chasse craignent plus la peste bovine que tout autre danger pouvant mettre en péril les animaux sauvages d’Afrique.
Osborn connaissait l’histoire de la peste bovine et il se livra à une étude des échanges internationaux entre espèces. Il était évident pour lui que si une seule maladie, venant d’une seule espèce allogène, pouvait prélever un tel tribut sur tout un continent, alors les pires catastrophes pouvaient survenir quand des faunes entières, des dizaines d’espèces venant de régions autrefois séparées, venaient à se mélanger. Le désastre serait inévitable. Dès lors qu’une espèce demeure dans une région biologique, elle s’adapte à ses prédateurs, à ses concurrents et à ses parasites. On appelle cela la loi de la coévolution. Sur une large échelle temporelle, la coévolution œuvrant sur des millions d’années permettra à tout un écosystème de s’équilibrer de toutes les manières possibles. Mais quand deux continents mélangent leur faune, chaque groupe se trouve confronté à de nouveaux ennemis contre lesquels il n’y a eu aucune préparation coévolutive.
Les germes et les bactéries ne sont pas les seuls touristes à apporter la mort. Des animaux plus grands peuvent jouer le même rôle. Un New-yorkais bien intentionné importa, il y a de nombreuses années, des étourneaux européens qu’il relâcha à Central Park pour permettre à Manhattan de jouir des oiseaux de la littérature anglaise. Les étourneaux ne sont pas, en principe, des nuisances dans leurs habitats de l’Ancien Monde. Mais en Amérique du Nord, ils déferlèrent comme des criquets emplumés ; aucune espèce locale ne put les arrêter. Les lapins ne posent pas de problèmes en Grande-Bretagne, mais lâchés en Australie, qui a développé une faune bien différente de celle de l’Europe, ils explosèrent car rien ne les contrôlait. Pendant presque tout l’Âge des Mammifères, l’Amérique du Sud était une île. Les mammifères et les oiseaux d’Amérique du Sud évoluèrent en une multitude d’espèces que l’on ne trouvait nulle part ailleurs (paresseux terrestres géants de six mètres de long, mammifères marsupiaux à dents de sabre, oiseaux aptères prédateurs plus gros que des lions, etc.). Les mammifères d’Amérique du Nord passèrent en Amérique du Sud voici deux millions d’années seulement quand l’isthme de Panama se forma. Parmi les immigrants, il y avait des représentants des éléphants, des jaguars, des cerfs, des tapirs, des loups, pour n’en nommer que quelques-uns. Ces immigrants d’Amérique du Nord dévastèrent la faune locale. La plupart des grandes espèces sud-américaines s’éteignirent, victimes des prédateurs et des concurrents venus du nord et de leurs maladies.
Le monde de la fin du Crétacé contenait les prémisses d’un tel désastre. Les mers peu profondes se retirèrent et une série d’extinctions balaya le monde marin. Une monumentale vague de dinosaures asiatiques émigra en Amérique du Nord tandis qu’une vague toute aussi importante de dinosaures américains envahit l’Asie. Dans toutes les régions touchées par ce mélange, des désastres, grands et petits, ont du se produire. Un prédateur étranger pouvait parfaitement réussir et ne pas être contrôlé, faisant un carnage de proies virtuellement sans défense tandis que ses populations atteignaient un niveau bien supérieur à ce qu’elles étaient dans son habitat naturel. Puis le prédateur pouvait brutalement disparaître, victime d’une maladie contre laquelle il n’était pas immunisé. Au fur et à mesure que les espèces se mélangeaient, venues de tous les coins du globe, le résultat ne pouvait être qu’un chaos biogéographique.
Un tel scenario n’est pas hypothétique et il ne nécessite pas une intervention extraterrestre. Un désastre global fut simplement le résultat inévitable de maladies et de nuisances lâchées à la fois sur les autochtones et les étrangers. Les pires effets étaient réservés aux plus grands voyageurs. Les grands animaux terrestres traversaient les barrières géographiques sans problèmes ; ils causaient plus de ravages et en subissaient plus. Les petites espèces ne peuvent pas migrer si facilement, parce que même une petite rivière pouvait arrêter leur progression. Ainsi l’extinction causée par le mélange des faunes devait être plus grave pour les animaux les plus grands et les plus actifs ; ce qui correspond exactement aux grandes extinctions de l’histoire géologique.
Comment l’endothermie colle-t-elle avec cette explication ? Le risque des dinosaures était-il supérieur s’ils possédaient un métabolisme élevé que s’ils étaient des vrais reptiles ? La réponse encore une fois est oui. Les reptiles a sang froid, à cause de leur métabolisme faible, voyagent mal. La quantité relativement faible d’énergie dont ils disposent les confine à de petits territoires et les oblige a une faible expansion géographique. Seuls les grands dépensiers en énergie ont besoin de grands territoires et repoussent en permanence leurs limites géographiques. Les vertébrés terrestres qui eurent une expansion rapide devaient être aussi les plus grands et métaboliquement les plus actifs. L’endothermie ajoute encore une dimension à leur vulnérabilité. Le meilleur moyen de nourrir une grande quantité de parasites est de les garder au chaud. Un serpent à sonnettes dans le désert décourage une telle incubation parce que sa température varie de 33 °C à presque 0 °C dans l’espace d’une journée. Mais du tissu gardé bien chaud par un métabolisme élevé doit être idéal du point de vue du parasite. Aujourd’hui, les animaux avec des températures corporelles constantes et élevées ont un cortège de parasites bien plus important que les reptiles et les amphibiens. Les dinosaures avec un métabolisme élevé risquaient bien plus l’extinction de masse que s’ils avaient été des reptiles géants avec un métabolisme faible.
Un tel scenario explique en détail l’extinction de masse sans avoir besoin de causes extraterrestres. La banalité de l’hypothèse d’Osborn est étonnante pour ceux qui sont fascinés par l’hypothèse de la collision céleste. Peut-être y a-t-il une place pour qu’un éclair céleste frappe les derniers survivants d’un écosystème affaibli. Mais la part la plus importante du crédit (ou du blâme) concernant le rythme de l’extinction et du refleurissement des espèces, sur terre comme sur mer, doit être porté au compte de la Terre elle-même, de son rythme et de ses conséquences biogéographiques naturelles.
NOTE
Il est fondamental d’être précis quand on parle de "diversité des dinosaures" dans les débats sur la fin du Crétacé. Depuis 1977, j’ai affirmé que le nombre d’espèces de grands dinosaures avait décliné bien avant le début de cette période. D. Russell prétend que le nombre total de toutes les espèces, n’a pas beaucoup changé. Je crois qu’on ne peut douter du déclin des espèces pesant plus d’une tonne. Triceratops a surclassé tous les autres genres dans presque tous les habitats durant les deux derniers millions d’années. Argumenter en partant du nombre total de petits dinosaures est prématuré car nous n’avons pas assez de crânes et de squelettes et une statistique basée sur des dents isolées, défavoriserait les prédateurs : les carnivores sont plus faciles à déterminer que les herbivores à partir de dents isolées.
Bibliographie
ALVAREZ, Walter, ASARO, Frank, MICHEL, Helen V. et ALVAREZ, Luis, "Evidence for a Major Meteorite Impact on the Earth 34 Million Years Ago : Implications for Eocene Extinctions", Science 216 (1982).
BAKKER, R.T., "Tetrapod Mass Extinctions- A Model of the Regulation of Speciation Rates and Immigration by Cycles of Topographic Diversity", in HALLAM, A., ed.-, Patterns of Evolution (Amsterdam, Elsevier Scientific Publishing Co, 1977), 439-468.
CROSBY, Alfred W., The Columbian Exchange : Biological and Social Consequences of 1492 (Westport, Greenwood Press, .1972).
ELTON, Charles S., The Ecology of Invasions by Animals and Plants (lère édition, London, Methuen 1958; 2ème éd. London, Chapmaniand Hall, 1972).
GANAPATHY, R., "A Major Meteorite Impact on the Earth 65 Million Years Ago : Evidence from the Cretaceous-Tertiary Boundary Clay", Science 209 (1980).
GOULD, Stephen Jay, "The Belt of An Asteroid", Natural History 89 (1980).
GOTTFRIED, Robert S., The Black Death : Natural and Human Disaster in Medieval Europe (New York, The Free Press, 1983).
KRASSILOV, V.A., "Climatic Changes in Eastern Asia as Indicated by Fossil Floras. 1. Early Cretaceous", Paleog. Paleocl. Paleoec. 13 (1973) : 261-274.
MacARTHUR, Robert H., Geographical Ecology : Patterns in the Distribution of Species (New York, Harper and Row, 1972).
McNEILL, William H., Plagues and Peoples (Garden City, Doubleday, 1976).
OSBORN, Henry Fairfield, The Age of Mammals in Europe, Asia and North America (New York, Macmillan, 1910).
RUSSELL, Dale A., "The Gradual Decline of the Dinosaurs -Fact or Fallacy?", Nature 307 (26) janvier 1984 : 360-361.
SIMPSON, George Gaylord, The Geography of Evolution (Philadelphie et New-York, Chilton Books, 1965).
ZINSSER, Hans, Rats, Mice and History (Boston, Little Brown and Cie,1934).